
In the early 1930s, Australian farmers were facing a big, feathery problem. Come wintertime, a migrating swarm of 20,000 large flightless birds known as emus would descend upon the Western Australian coast, ravaging farmland along the way.1 Reaching up to 6 feet tall and weighing 130 pounds, an adult emu is no pushover. But the Australian government would not take these bird-brained pests lying down. In November of 1932, the Royal Australian Artillery was deployed to deal with the fowl, whereupon they quickly realized that the emus’ evasive behavior and surprising durability made them poor targets for machine gun fire.2 Thus, the so-called Great Emu War ended with a swift Aussie surrender, and the emu menace would continue to terrorize Australian farmers for many years.
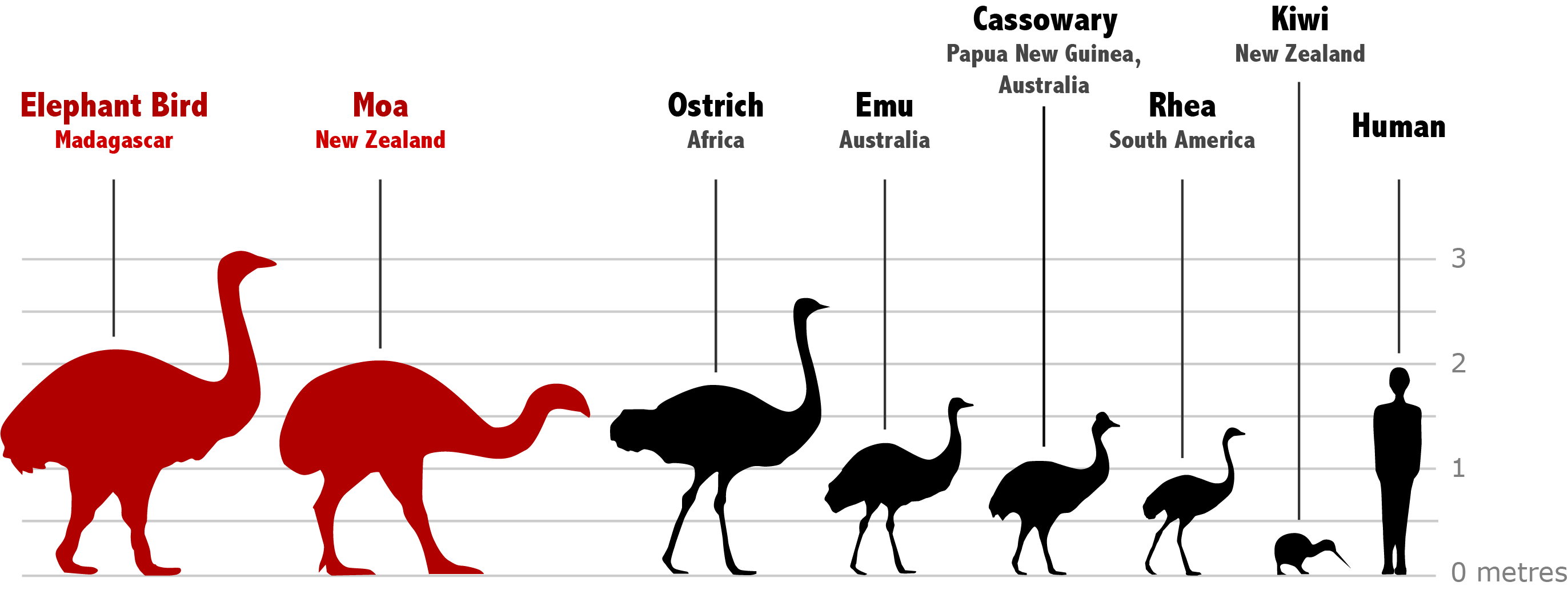
Unfortunately, the largest flightless birds did not fare as well as emus in the face of human persecution. Both the moas of New Zealand and the elephant birds of Madagascar were wiped out following human inhabitation of their respective islands.3,4 As a consequence, ostriches are the largest birds alive today, followed by the cassowaries of New Guinea and Australia. Other giant flightless birds include the aforementioned emu and the rheas of South America. Together with the comparatively miniscule kiwis, these land-loving birds form the group known as the ratites (Figure 1, Figure 2). The ratites are distinguished—even from other flightless birds—by their lack of a keel, a protrusion of the breastbone that acts as an anchor for the muscles used in powered flight.5 Despite being so utterly earthbound, these birds are found in disparate islands and continents around the world, giving rise to a compelling mystery as to their method of distribution.
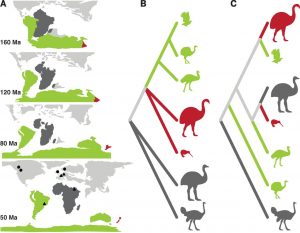
In addition to their lack of a keel, the ratites share a plethora of anatomical features that indicate a shared evolutionary history.5,6 Therefore, the intuitive and classical view has been that this group of birds descended from a flightless ancestor living on Gondwana, an ancient supercontinent which encompassed modern-day Africa, South America, and Australia. This postulate, referred to as the vicariance hypothesis, asserted that ratites were spread across the Southern Hemisphere as the continents composing Gondwana separated over the course of the past 180 million years (Figure 3A, B).5 Recent evidence, however, suggests that although these birds are indeed related, they evolved their large, flightless bodies independently. This would make the ratites an example of “convergent evolution,” where multiple species develop a shared set of features in response to similar conditions. For a classic example of convergent evolution, think of the similar torpedo-shaped forms of dolphins and sharks. Another widespread example are crabs—whose flat, side-scuttling body plan has famously been adopted by crustaceans so many times that the process has a dedicated word: carcinization.7

Birds of a Feather Didn’t Stop Flying Together
All life on Earth, from the smallest bacterium to the beefiest ostrich, is unified by the genetic code—a system by which the instructions for life are stored within DNA and passed down through countless generations. Errors or damage to DNA can induce small changes, known as mutations. Although the word “mutation” often carries a negative connotation, most of these changes are actually harmless. Thus, these mutations accumulate, and the stockpile of distinct mutations in one lineage can be used to distinguish it from another. On a small scale, the mutations you share with your siblings would help somebody analyzing your DNA determine genetic relevancetell that you’re related. Likewise, species who last shared a common ancestor millions of years ago can have their evolutionary family trees pieced together through tracking their mutations.8
In addition, by counting the number of mutations separating two lineages, and with knowledge of the rate at which such mutations occur over time, it is possible to estimate how much time has passed since two groups diverged. This idea—that the number of mutations can be used to calculate the time since evolutionary divergence—is known as the molecular clock, a powerful tool used to unveil evolutionary history, especially in combination with the fossil record.9
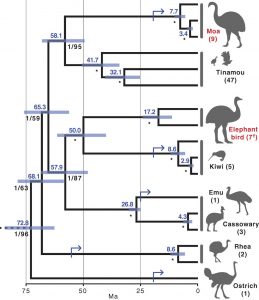
Application of these genetic tools to the ratites has yielded startling results, with relationships and divergence times that contradict the vicariance hypothesis (Figure 3C, Figure 4). For example, the closest relatives of the diminutive kiwis were the recently extinct enormous elephant birds of Madagascar, and these groups appear to have diverged after their landmasses were separated.10,11 Most damning for the vicariance hypothesis is the placement of the flighted South American birds known as tinamous within the ratite clade. As it turns out, tinamous are the closest relatives of the now-extinct—and very flightless—moas of New Zealand.10, 11, 12, 13 In contrast, the ostrich diverged from other ratites 70 to 80 million years ago, well before the moa could have figuratively snipped its wings. In other words, the small, flight-capable tinamou is sister to moas, elephant birds, and emus while the ostrich is a cousin. The basal ratites—that is, their figurative grandparents—must therefore have been volant (meaning flight capable). Based on skeletal structure, a likely identity for these ancestors is the group of fossil birds called the lithornithids, whose oldest fossils are found in Eurasia.11, 14 Thus, the most compelling, current argument for the evolution of the ratites is that they originated as small volant birds in the Northern Hemisphere before spreading to the Southern landmasses of former Gondwana, where they lost flight—and gained size—independently of one another.10, 12, 13 What extraordinary turn of events could have led all of these birds to forgo a life in the sky?
It’s Free Real Estate
Sixty-six million years ago, a meteor with the mass of at least 10 trillion emus (0.3 to 3 quadrillion kilograms) struck the Gulf of Mexico, initiating a catastrophic global extinction event that wiped out the non-avian dinosaurs.15 With these dominant large land animals gone, birds had free reign to take over the void they left behind. One study theorizes that loss of flight in multiple ratite lineages occurred soon after this Cretaceous-Tertiary (K-T) extinction event (Figure 5).13 As the ratites evolved large body sizes to fill the large, herbivorous roles once held by the dinosaurs, flight became a hindrance. The increase in the energy cost of flight outpaces the increase in muscle power provided by a larger body.13 Combined with the fact that the ratites no longer needed to fear their previous land-based predators, evolutionary pressures may have pushed them to give up the ability to fly for greater size. The exception to this is the kiwi, the most omnivorous and smallest of the ratites. It is possible that, upon arriving to New Zealand, the ancestor of the kiwi found the large herbivore niche already occupied by the moas. Kiwis, therefore, shed their flight feathers for another reason: living on an island safe from predators.
Loss of flight among island birds is actually relatively common. For example, up to a third of bird species in New Zealand before human habitation may have been flightless, and a group of birds known as rails has become flightless dozens of times in the process of colonizing scattered islands across the world’s oceans.16, 17 An island habitat free of mammalian predators—such as New Zealand—renders many of the advantages of flight moot. Because of how costly flight is to maintain—flight muscles demand a minimum of 12% of a bird’s body mass, and up to 40% in extreme cases—the ability to fly is quickly lost if it is no longer advantageous.16
In addition, island habitation may have contributed to the abnormal size of some of the ratites. The phenomenon of island gigantism refers to the trend of animals on island habitats evolving to be much larger than their mainland counterparts.18 In the same manner that islands remove the need for flight, they remove constraints on size. Small herbivores, for example, may no longer need to hide from predators. This pattern might explain why the elephant birds and moas were so large, even in comparison to other ratites. However, this theory can be contentious, and it is possible that elephant bird and moa gigantism was a response to global cooling.19
Because of one or both of these reasons, the ancestors of the ratites lost the ability to fly anywhere from four to six separate times.13 This means that the similar body structure of the ratites is a product of convergent evolution. It seems that having a long neck and gangly legs is somehow an ideal physiology for a large, flightless bird.

Look at All Those Emus
While some people work to keep these birds at bay, others are busy spreading them across the world. In the most recent 2017 census, there were over 11,500 emus, 4,700 ostriches, and 1,300 rheas being raised by farms within the United States, where they are grown for meat and eggs—which are up to twenty times the weight of a chicken’s.20, 21 In Germany, seven greater rheas managed to escape from such a farm in the late 1990s, and their population has since ballooned into the hundreds. Echoing their Australian cousins, these wild rheas in Germany have become an agricultural pest. Rather than declaring war, however, the German government attempts to handle the rhea by legalizing hunting.22 Time will tell if these efforts prove successful, but these birds truly should not be underestimated. Maybe when you’re the closest thing to a living, breathing, dinosaur, you really don’t need to fly.
References
- ‘Feathered foes’: Soldier settlers and Western Australia’s ‘Emu War’ of 1932. (n.d.). https://doi.org/10.1080/14443050609388083
- New Strategy In A War On The Emu. (1953, July 5). Sunday Herald. http://nla.gov.au/nla.news-article18516559
- Crowley, B. E. (2010). A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quaternary Science Reviews, 29(19), 2591–2603. https://doi.org/10.1016/j.quascirev.2010.06.030
- Holdaway, R. N., Allentoft, M. E., Jacomb, C., Oskam, C. L., Beavan, N. R., & Bunce, M. (2014). An extremely low-density human population exterminated New Zealand moa. Nature Communications, 5(1), Article 1. https://doi.org/10.1038/ncomms6436
- Cracraft, J. (1974). Phylogeny and Evolution of the Ratite Birds. Ibis, 116(4), 494–521. https://doi.org/10.1111/j.1474-919X.1974.tb07648.x
- Fowler, M. E. (1991). Comparative Clinical Anatomy of Ratites. Journal of Zoo and Wildlife Medicine, 22(2), 204–227.
- Keiler, J., Wirkner, C. S., & Richter, S. (2017). One hundred years of carcinization – the evolution of the crab-like habitus in Anomura (Arthropoda: Crustacea). Biological Journal of the Linnean Society, 121(1), 200–222. https://doi.org/10.1093/biolinnean/blw031
- Kasha, M. (1962). Horizons in biochemistry. Albert Szent-Györgyi dedicatory volume. New York, Academic Press. http://archive.org/details/horizonsinbioche0000kash
- Donoghue, P. C. J., & Yang, Z. (2016). The evolution of methods for establishing evolutionary timescales. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1699), 20160020. https://doi.org/10.1098/rstb.2016.0020
- Mitchell, K. J., Llamas, B., Soubrier, J., Rawlence, N. J., Worthy, T. H., Wood, J., Lee, M. S. Y., & Cooper, A. (2014). Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science, 344(6186), 898–900. https://doi.org/10.1126/science.1251981
- Yonezawa, T., Segawa, T., Mori, H., Campos, P. F., Hongoh, Y., Endo, H., Akiyoshi, A., Kohno, N., Nishida, S., Wu, J., Jin, H., Adachi, J., Kishino, H., Kurokawa, K., Nogi, Y., Tanabe, H., Mukoyama, H., Yoshida, K., Rasoamiaramanana, A., … Hasegawa, M. (2017). Phylogenomics and Morphology of Extinct Paleognaths Reveal the Origin and Evolution of the Ratites. Current Biology, 27(1), 68–77. https://doi.org/10.1016/j.cub.2016.10.029
- Smith, J. V., Braun, E. L., & Kimball, R. T. (2013). Ratite Nonmonophyly: Independent Evidence from 40 Novel Loci. Systematic Biology, 62(1), 35–49. https://doi.org/10.1093/sysbio/sys067
- Phillips, M. J., Gibb, G. C., Crimp, E. A., & Penny, D. (2010). Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites. Systematic Biology, 59(1), 90–107. https://doi.org/10.1093/sysbio/syp079
- Leonard, L. M., Dyke, G. J., & Tuinen, M. van. (2005). A new specimen of the fossil palaeognath Lithornis from the Lower Eocene of Denmark. American Museum novitates ; no. 3491. https://digitallibrary.amnh.org/handle/2246/5660
- Pope, K. O., Baines, K. H., Ocampo, A. C., & Ivanov, B. A. (1997). Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact. Journal of Geophysical Research: Planets, 102(E9), 21645–21664. https://doi.org/10.1029/97JE01743
- McNab, B. K. (1994). Energy Conservation and the Evolution of Flightlessness in Birds. The American Naturalist, 144(4), 628–642.
- Kirchman, J. J. (2012). Speciation of Flightless Rails on Islands: A DNA-Based Phylogeny of the Typical Rails of the Pacific. The Auk, 129(1), 56–69. https://doi.org/10.1525/auk.2011.11096
- Lomolino, M. V. (2005). Body size evolution in insular vertebrates: Generality of the island rule. Journal of Biogeography, 32(10), 1683–1699. https://doi.org/10.1111/j.1365-2699.2005.01314.x
- Crouch, N. M. A., & Clarke, J. A. (2019). Body size evolution in palaeognath birds is consistent with Neogene cooling-linked gigantism. Palaeogeography, Palaeoclimatology, Palaeoecology, 532, 109224. https://doi.org/10.1016/j.palaeo.2019.05.046
- United States. (2017) U.S. Census of Agriculture. United States. Retrieved from https://www.nass.usda.gov/Publications/AgCensus/2017/
- Cameron, A., Perrins, C. M., Harrison, C. J. O., & Reader’s Digest Association. (1979). Birds—Their life, their ways, their world. Pleasantville, N.Y. : Reader’s Digest Association. http://archive.org/details/birdstheirlifeth00came
- Inside Germany’s Giant, Hungry, Flightless-Bird Problem. (2020, September 26). Audubon. https://www.audubon.org/news/inside-germanys-giant-hungry-flightless-bird-problem
Image References
Cover Image: Quine, T. (2015). American Museum of Natural History, New York City, 2015. Extinct flightless bird. https://commons.wikimedia.org/wiki/File:Extinct_flightless_bird_(21128386393).jpg
.jpg){kind=link}
Figure 1: Palaeognathae. (2023). In Wikipedia. https://en.wikipedia.org/w/index.php?title=Palaeognathae&oldid=1145670875
Figure 2: Taonga, N. Z. M. for C. and H. T. M. (n.d.). Ratite birds [Web page]. Ministry for Culture and Heritage Te Manatu Taonga. Retrieved February 27, 2023, from https://teara.govt.nz/en/diagram/11361/ratite-birds
Figures 3 & 4: Mitchell, K. J., Llamas, B., Soubrier, J., Rawlence, N. J., Worthy, T. H., Wood, J., Lee, M. S. Y., & Cooper, A. (2014). Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science, 344(6186), 898–900. https://doi.org/10.1126/science.1251981
Figure 5: Phillips, M. J., Gibb, G. C., Crimp, E. A., & Penny, D. (2010). Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites. Systematic Biology, 59(1), 90–107. https://doi.org/10.1093/sysbio/syp079